
Showing posts from February 18, 2011Show all
 Zoology
Zoology
Metabolism of Fats and Proteins and Control of Metabolism and Metabolic Pool
Catabolism of glucose is most common metabolic pathway in cells. Animals are consumed…
 Zoology
Zoology
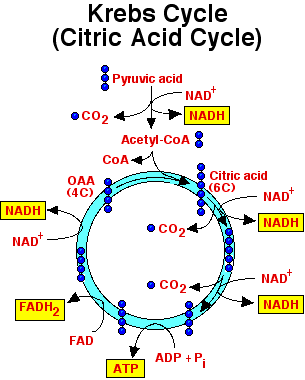
Kreb’s Cycle (OR) Citric Acid Cycle (OR) Tri-Carboxylic Acid Cycle (TCA)
During metabolism the synthesis and breakdown of different organic compounds takes pl…

Social Plugin